SNF1与
MetK1共表达促进酵母S-腺苷-L-甲硫氨酸合成
陈海龙, 戴佳良, 邓晶, 高新星, 朱官兴, 何清明, 朱年青
中国农业科技导报
2025, 27 ( 10):
84-94.
DOI:10.13304/j.nykjdb.2024.0747
S-腺苷-L-甲硫氨酸(S-adenosyl-L-methionine,SAM)是生物体内重要的生理活性物质。通过调控酿酒酵母的葡萄糖效应而将碳通量定向到SAM合成是细胞工厂的挑战。以酿酒酵母CGMCC 2842(2842)为出发菌,过表达SNF1基因调控葡萄糖效应并引入利士曼原虫(Leishmania infantum)SAM合成酶基因MetK1。结果发现,SNF1过表达可提高糖酵解相关基因表达水平和葡萄糖利用率,进而改善糖酵解中间体水平;提高ACS1、ALD6表达及乙醇脱氢酶2(alcoholdehy drogenase 2,ADH2)活性,可促进发酵液中的乙醇转化为乙酰辅酶A;还增强了SAM前体氨基酸及含硫氨基酸代谢相关基因表达水平。异源MetK1引入可有效定向碳代谢到SAM合成。重组菌YPSNF1-MetK1菌株的SAM产量达1.90 g·L-1,较出发菌株2842产量提高251.85%。以上研究结果为酵母葡萄糖效应调控及其应用奠定理论基础。
| 基因Gene | 正向引物Forward primer (5’-3’) | 反向引物Reverse primer (5’-3’) |
|---|
| HXT1 | TGTGCCATTGGTGGTATCGT | ACCACCGACACCTAAACCAG | | HXK1 | TACTGGTGTCAACGGTGCTT | GTTCGTCGACAGCAACATCG | | HXK2 | CTGCTCCAATGGCCATCAAC | AAGGTTTGTTGGCCTGGTCT | | PFK1 | TGGTCTTGTCGGTTCCATCG | AAGGTTTGTTGGCCTGGTCT | | TDH1 | TTGAGGTTGTTGCTGTCAACG | GCTTGTCGTCATGGGAAACAG | | PGK1 | AGGCTTCTGCCCCAGGTTC | CAGCACGTTGTGGCAAGTC | | PYK | CGACTCAGATGCTGGATTCA | CCGTTTCTCCAGAAAGCATAA | | ALD6 | GAACTTCACCACCTTAGAGCCA | GCAGCGGGTTTCAAGATACA | | ACS1 | GCTGCAAAGGATAAGGATGG | GCCTCAATTTCAGCGGTAGA | | SER33 | CTCGTCGTGTAAGCATT | ACTTGTGGAACTCTACTT | | SER1 | ACAACTCAGCCTATAATACAA | CCAATGCCTCATATAATATCTT | | SER2 | CCTATCGTAGACGGACAG | CACCATACAACTTGCTTCA | | MET17 | AAATGGATTGGTGGTCATGG | GAAAGAGGCAAATGGGTTCA | | MET6 | TGGAAGCTGCCGGTATCAAG | GCAACTCTGAAAGCTTCGGC | | SAM2 | TGAATCCGTCGGTGAAGGTC | AGCTGTTTCACAGGCAACCT | | ACT1 | ACGCTCCTCGTGCTGTCTTC | GTTCTTCTGGGGCAACTCTCA |
View table in article
表1
RT-qPCR 分析所用引物
正文中引用本图/表的段落
S-腺苷-L-甲硫氨酸(S-adenosyl-L-methionine,SAM)是生物体内重要的生理活性物质。通过调控酿酒酵母的葡萄糖效应而将碳通量定向到SAM合成是细胞工厂的挑战。以酿酒酵母CGMCC 2842(2842)为出发菌,过表达SNF1基因调控葡萄糖效应并引入利士曼原虫(Leishmania infantum)SAM合成酶基因MetK1。结果发现,SNF1过表达可提高糖酵解相关基因表达水平和葡萄糖利用率,进而改善糖酵解中间体水平;提高ACS1、ALD6表达及乙醇脱氢酶2(alcoholdehy drogenase 2,ADH2)活性,可促进发酵液中的乙醇转化为乙酰辅酶A;还增强了SAM前体氨基酸及含硫氨基酸代谢相关基因表达水平。异源MetK1引入可有效定向碳代谢到SAM合成。重组菌YPSNF1-MetK1菌株的SAM产量达1.90 g·L-1,较出发菌株2842产量提高251.85%。以上研究结果为酵母葡萄糖效应调控及其应用奠定理论基础。
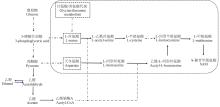
S-腺苷-L-甲硫氨酸(S-adenosyl-L-methionine,SAM)在人体内作为甲基、巯基和氨丙基供体参与代谢反应、调节基因表达和信号转导等,常被用于肝脏、骨关节炎和抑郁症等疾病的临床防治[1-3]。其在细胞内的代谢途径如图1所示。SAM在调控植物对缺铁[4]、干旱[5]和盐碱[6]的适应性以及在抵抗病原菌[7]和衰老进程[8]中也发挥重要作用。刘鑫等[9]发现,外源SAM能显著促进植物扦插苗根系生长发育、叶绿素合成,并能增强植物对氮、磷、钾等元素的吸收,SAM浸泡处理还可增加黄瓜下胚轴生长素和多胺的合成及其相关基因表达水平。
基于此,本研究以酿酒酵母CGMCC 2842为出发菌株,构建
SNF1过表达重组酵母菌株,并比较利士曼原虫(
Leishmania infantum)的SAM合成酶基因
MetK1和酵母的
SAM2基因促进SAM合成的能力,最终构建
SNF1和
MetK1共表达重组酵母菌株,并在含有不同葡萄糖含量(5%和10%)的发酵培养基中发酵,考察重组菌胞内SAM积累量、葡萄糖消耗情况、生物量、代谢中间产物、前体氨基酸及乙醇代谢等发酵参数的变化情况,分析
SNF1和
MetK1对目标产物SAM在酿酒酵母胞内的合成与积累的影响,以期获得稳定高产SAM重组菌,为酿酒酵母葡萄糖效应的调控以及SAM规模化生产奠定基础。
酿酒酵母CGMCC 2842(简称2842)和酿酒酵母表达质粒pYES-KanMx为中国药科大学周长林教授课题组惠赠。重组表达载体pYES-SNF1、pYES-SAM2、pYES-MetK1、pGAL-SNF1-pPGK-MetK1及重组酵母菌YPGK、YPSAM2、YPSNF1、YPMetK1、YPSNF1-MetK1为本研究构建。
将
SNF1和酵母表达质粒pYES-KanMx分别用
Hind Ⅲ和
BamH Ⅰ酶切,T4 DNA连接酶连接,采用CaCl
2法转化
Escherichia coli DH5α感受态细胞,涂布在含有100 μg·mL-1氨苄青霉素的LB培养基平板上,37 ℃培养过夜,挑取阳性克隆于5 mL LB培养基过夜培养,用SanPrep柱式DNA质粒提取试剂盒〔生工生物工程(上海)股份有限公司〕提取重组质粒,采用0.8%琼脂糖凝胶电泳检测。重组质粒用
Hind Ⅲ和
BamH Ⅰ双酶切并测序验证后,获得重组表达质粒pYES-SNF1。其他重组质粒构建方法相同,pYES-SAM2和pYES-MetK1构建时采用
Xho Ⅰ和
Sph Ⅰ酶切位点。
SNF1和
MetK1共表达质粒pGAL-SNF1-pPGK1-MetK1构建时,
SNF1插入位点采用
Hind Ⅲ和
BamH Ⅰ酶切,启动子
pPGK1引入采用
EcoR Ⅰ和
Not Ⅰ酶切,
MetK1插入采用
Xho Ⅰ和
Sph Ⅰ酶切。
糖酵解和前体氨基酸代谢相关基因表达水平测定。利用酵母总RNA分离试剂盒[生工生物工程(上海)股份有限公司]从酵母细胞中提取总RNA。通过1%琼脂糖凝胶验证RNA质量,并通过核酸蛋白测定仪(艾本德中国有限公司)定量。按照PrimeScript RT试剂盒[宝生物工程(大连)有限公司]说明书进行反转录。定量分析靶基因
HXT1(编码己糖转运蛋白,HXT1)、
HXK1(编码己糖激酶1,HXK1)、
HXK2(编码己糖激酶2,HXK2)、
PFK1(编码6-磷酸果糖激酶1,PFK1)、
TDH1(编码甘油醛-3-磷酸脱氢酶,TDH1)、
PGK1(编码3-磷酸甘油酸激酶,PGK1)、
PYK(编码丙酮酸激酶,PYK)、
ALD6(编码乙醛脱氢酶6,ALD6)和
ACS1(乙酰辅酶A合成酶1)、
SER33(编码磷酸甘油酸脱氢酶,SER33)、
SER1(编码磷酸丝氨酸转氨酶,SER1)、
SER2(编码磷酸丝氨酸磷酸酶,SER2)、
MET17(编码o-乙酰-高丝氨酸巯基化酶,MET17)、
MET6(编码蛋氨酸合成酶,MET6)、
SAM2(编码SAM合成酶2,SAM2)和内参基因
ACT1的引物序列见表1。使用荧光定量PCR仪(ABI)对上述靶基因进行定量[30?31]。
由图2可知,5%葡萄糖中YPSNF1、YPSAM2、YPMetK1和YPSNF1-MetK1菌株的SAM积累量分别为0.76、1.18、1.41和1.66 g·L-1,较出发菌株2842(0.60 g·L-1)分别提高26.7%、96.6%、135.0%和177.0%;而在10%葡萄糖中,YPSNF1、YPSAM2、YPMetK1和YPSNF1-MetK1菌株的SAM积累量分别为0.90、0.88、1.14和1.90 g·L-1,较出发菌株2842分别提高66.7%、63.0%、111.0%和251.9%。其中,YPSNF1菌株的SAM积累量均显著高于出发菌株,表明
SNF1过表达可以显著改善酿酒酵母胞内SAM的合成与积累,在高葡萄糖中更明显;另外,YPMetK1菌株的SAM产量显著高于YPSAM2,表明引入利士曼原虫的
MetK1较同源
SAM2能更显著改善SAM产量。
测定2842、YPSNF1、YPMetK1和YPSNF1-MetK1菌株的生长曲线和葡萄糖消耗曲线,结果(图3)发现,在5%和10%葡萄糖中,YPSNF1、YPSNF1-MetK1的细胞干重较出发菌株2842显著提高,而YPMetK1变化不显著(图3A和B),表明
SNF1过表达有利于生物量积累;同时发现YPSNF1、YPMetK1、YPSNF1-MetK1较出发菌株2842具有更强的葡萄糖消耗能力,尤其是
SNF1的过表达显著改善葡萄糖利用情况,在10%葡萄糖中表现更为显著(图3C和D)。
为进一步展示葡萄糖利用的提高,计算10%葡萄糖处理中目标产物SAM对葡萄糖和细胞干重(DCW)的得率,结果(表2)发现,YPMetK1、YPSNF1和YPSNF1-MetK1菌株SAM对DCW得率分别为121.95、136.04和229.75 mg·g-1,较出发菌株2842分别提高68.3%、87.6%和216.8%;其SAM对葡萄糖得率分别为9.00、11.40和19.00 mg·g-1,较出发菌株2842分别提高68.3%、87.6%和216.8%。由此表明,
SNF1的过表达和
MetK1的引入都显著改善了目标产物SAM对葡萄糖及DCW的得率,即
SNF1和
MetK1的过表达提高了酵母对葡萄糖的利用率。
为探究上述重组菌葡萄糖利用率提高的原因,考察重组菌糖酵解代谢途径中间代谢产物积累水平和相关基因表达水平的变化,结果(图4A)发现,在5%葡萄糖中,YPSNF1和YPSNF1-MetK1胞内G6P水平分别为0.32和0.28 g·L-1,较出发菌株2842分别提高45.5%和27.3%;YPSNF1和YPSNF1-MetK1胞内FBP水平分别为0.43和0.37 g·L-1,较出发菌株2842菌株分别提高95.5%和68.2%,且胞内PY水平较出发菌株2842分别增长88.9%和61.1%,乙酰辅酶A水平分别提高34.4%和18.8%。同样的现象在10%葡萄糖中更为显著(图4B)。YPMetK1菌株在5%葡萄糖中的G6P、FBP、PY和乙酰辅酶A水平较出发菌株2842显著降低,表明
MetK1的引入有利于这些代谢中间产物定向到SAM合成;YPMetK1菌株在10%葡萄糖中,上述中间产物较出发菌株2842有所下降,但差异不显著,可能与葡萄糖水平较高有关。
由图4C和D可知,虽然YPSNF1、YPSNF1-MetK1菌株的
PYK和
ALD6相对表达水平在5%葡萄糖中差异不显著,但在5%和10%葡萄糖组中,YPSNF1、YPSNF1-MetK1菌株
HXT1、
HXK1、
HXK2、
PFK1、
TDH1、
PGK1、
PYK、
ALD6和
ACS1基因的相对表达量较2842均显著提高,尤其是在10%葡萄糖中,在YPSNF和YPSNF1-MetK1菌株中上述基因的表达水平增加更为显著。然而与5%葡萄糖组相比,YPMetK1菌株在10%葡萄糖中相关基因表达水平并未呈现出显著的提高,除
TDH1和
ACS1基因外,其他基因表达水平均下降(图4C和D)。由此表明,
SNF1的过表达能够显著提高酿酒酵母糖酵解相关基因的表达水平,进而显著提高细胞内糖酵解中间体水平,而
MetK1的引入有利于这些代谢中间产物定向到SAM合成。
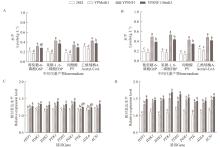
对这些氨基酸代谢相关的基因进行定量分析,结果(图5C和D)发现,除
SER1基因外,YPSNF1和YPSNF1-MetK1在5%葡萄糖中胞内的
SER33、
SER2、
MET17、
MET6和
SAM2基因转录水平较2842显著上调;而在10%葡萄糖中YPSNF1和YPSNF1-MetK1胞内的
SER33、
SER1、
SER2、
MET17、
MET6和
SAM2基因的转录水平较2842上调更显著。
SER33、
SER1和
SER2共同催化G3P转化为Ser,这是含硫氨基酸生物合成的关键节点。作为
SER33、
SER1、
SER2、
MET17、
MET6和
SAM2转录水平上调的结果,细胞内Ser、Asp、Cys和Met等SAM前体氨基酸在胞内的水平也增加。由此表明,
SNF1过表达显著上调
SER33、
SER1、
SER2、
MET17、
MET6和
SAM2等相关基因转录水平,有利于酿酒酵母胞内Cys、Asp、Ser和Met的合成与积累;而
MetK1的引入则可以有效地将Cys、Asp、Ser和Met等前体氨基酸定向到SAM合成。
综上表明,过表达
SNF1可以提高酿酒酵母中负责催化乙醇氧化反应生成乙酰辅酶A的ADH2活性及
ALD6、
ACS1基因的表达水平,进而降低发酵液中乙醇的积累;乙醇积累量的降低意味着可能有更多的碳源转向SAM或其他代谢物的合成和酵母的生长。
研究表明,
SNF1过表达可增强酵母对环境胁迫的响应,并改善葡萄糖利用[30,37?38]。另外,在酵母细胞乙醇脱氢酶中,ADH1、ADH3、ADH4和ADH5蛋白负责将乙醛还原为乙醇,而ADH2负责将乙醇氧化成乙醛。ADH2和6型乙醛脱氢酶(ALD6,由
ALD6基因编码)、1型乙酰辅酶A合成酶(ACS1,由
ACS1基因编码)协同催化乙醇转化为乙酰辅酶A[39?40]。当葡萄糖存在时,SNF1复合体抑制
ADH2表达;当葡萄糖耗尽时,SNF1复合体被激活,进而使得
ADH2解除抑制,表达出催化乙醇氧化的ADH2活性[39]。本研究发现,
SNF1过表达可提高葡萄糖运输和糖酵解相关基因表达水平,葡萄糖利用率得到改善,进而提高糖酵解中间体水平;特别是在葡萄糖过量存在时,
ACS1和
ALD6表达水平及ADH2活性增强,可促进发酵液中积累的乙醇转化为乙酰辅酶A;还增强了SAM前体氨基酸及含硫氨基酸代谢相关基因表达水平。综上表明,
SNF1过表达的多重效应对于提高酵母胞内SAM合成表现出很大潜力。
本研究通过过表达酿酒酵母葡萄糖效应调控相关基因
SNF1以实现减轻其葡萄糖效应,同时引入来自利士曼原虫SAM合成酶基因
MetK1,将碳代谢流定向到目标产物SAM的合成,以期获得稳定、高产的SAM重组菌,为酿酒酵母葡萄糖效应的调控以及SAM规模化生产奠定基础。
本文的其它图/表
|